![]()
CHAPTER 3 - VEGETATION STRUCTURE AS AN INDICATOR FOR LAND-COVER DYNAMICS ASSESSMENT IN THE AMAZON
3.5. The role of vegetation structure and remote sensing for the study of secondary succession dynamics in colonization areas of Amazônia
The study of vegetation structure presented in this chapter confirmed expected trends about secondary succession of tropical forests in Rondônia, which include: increase in density of trees with decrease in density of saplings; increase in DBH of trees and total basal area; increase in total height of trees; and, consequently, increase in total aboveground biomass. In addition, the results obtained for selected vegetation structure variables were depicted by spectral responses in Landsat TM bands, particularly the ones within the infrared portion of the spectrum. This last assertion has already been discussed in the literature and is due to chlorophyll absorption in the visible TM bands (1-3); mesophyll reflectance in the near infrared (band 4); and for both plant and soil water absorption in the mid-infrared bands (5 and 7) (Moran et al. 1994). The balance within and between these three groups of bands permits the differentiation of stages of succession, tropical forest, and other land-cover classes.
The applicability of these findings surpasses the understanding of vegetation recovery processes at local scales. It allows the spatial-temporal monitoring of Amazonian landscapes regarding their land-cover dynamics. Being able to differentiate distinct stages of vegetation regrowth in a landscape makes it possible to draw a better picture about LULC trajectories. The use of remote-sensing techniques has improved this capability by ensuring the investigation of secondary succession in larger areas on a multi-temporal basis. However, such an enterprise is not an easy task, mainly because it artificially reduces the continuous process of vegetation recovery to a selected number of categories.
The dichotomy between natural heterogeneity and the scientific need for generalizations is a major challenge in ecological research. As the knowledge about a phenomenon being studied increases and becomes available for the scientific community, interpolations and extrapolations are readily made. The scales of space, time, and complexity are then reduced to relatively few discrete explanatory categories (Wilson 1998). It is common to see broad generalizations based on local factors and/or grouping of local heterogeneity based on generalizations.
O'Neill et al. (1986), willing to justify such attitude, recognized that some degree of abstraction is required in order to study ecological systems. The rationale behind this statement is that ecology cannot set up a single spatial-temporal scale that will be adequate for all investigations. Space-time dynamics is now understood as a central issue related to ecosystem studies (Kareiva 1994). The scale factor, widely discussed in a variety of studies, has induced the development of a hierarchical view of ecosystems (Allen and Star 1982, Bian 1997). Instead of descriptive and qualitative attempts to analyze natural heterogeneity and dynamics, quantitative approaches have shown the development of complex ecological systems toward new levels of organization (Sklar and Costanza 1991).
These processes range from local to global scales, and the argument is valid for vegetation ecological studies. This chapter focuses on tropical forests in Rondônia and their stages of succession after disturbance. If there is a common sense about what forest is, it relates to a 'vegetation community dominated by trees.' However, even within specific biomes, they may differ in structure, composition, and physiognomy. From the ecologist's standpoint, forests consist of a mosaic of gap-phase, building-phase, and mature-phase formations (Whitmore 1998).
The tentative nature of defining secondary succession classes in Rondônia based on vegetation structure data and remote sensing complies with the need to monitor land cover in the Amazon. But how many final classes need to be defined and how do they correspond with the classes used during field sampling? Do TM spectral bands depict all classes identified by the original sample? If not, which variables can be used to control the process of categorization?
One way to address these questions is to follow the regrowth trajectory in selected sites and assign stages based on age (Uhl 1987, Guariguata et al. 1997, Nelson et al. 1999). This method generally maintains control over site variability but does not allow generalization to other areas within the region due to the small number of samples. In addition, the vegetation classes are difficult to depict in TM images at this level of separation. Another way to address the heterogeneity of vegetation classes of regrowth in the Amazon is to define a range of classes based on vegetation structure (Brondizio 1996, Tucker 1996) or age (Uhl et al. 1988, Steininger 1996). This method may also be affected by the sampling strategy. For instance, if age is considered as an initial parameter to define regrowth classes, gaps between ages sampled may produce additional lack of information in order to identify the final classes. However, using stratified random sample techniques minimizes the potential pitfalls of such strategy by ensuring a broader representation of the natural variability in the study area.
For the research in Rondônia, the sample did not include ages 1, 2, 4, 7, 11, and greater than 14 years. The decision of undersampling the first and second years of succession was discussed above. In this case, the confusion with pasture in terms of land use, vegetation structure (similar to degraded pasture), and spectral responses, indicated that the accuracy of assigning SS1 increases if stands more than 2 years old are considered. The decision was supported by previous studies showing that most class errors were associated with youngest age classes (< 2 years) and with different successional pathways and vegetation composition (Foody et al. 1996). To avoid the problem, Steininger (1996) also assigned classes of regrowth starting at two years old, and included a class called 'pasture with trees' between 'farmland' (agriculture and pasture) and secondary succession.
The confusion between SS1 and degraded pasture is mostly due to a common practice among local landowners. After slashing and burning, they often seed grass for pasture. If not, they plant annual crops, then seed grass. Or they plant annual and perennial crops (mostly coffee), and, if anything goes wrong, planting grass is again seen as an alternative land use. With the use of fire, grasses tend to overcome other pioneer species, playing an important role in initial stages of succession and consequently affecting spectral classifications. After the second year, recovery by saplings and small trees diminish the importance of grasses regarding ground cover. Then, the physiognomy (structure) of a fallow is better characterized.
On the other hand, not including first- and second-year SS1 increased the averages for some key structural variables being analyzed. For example, total height and DBH of trees for SS1 in the study area were greater than reported by other comparable studies (Alves et al. 1997). Besides this initial period of regrowth and a few years of not being present in the sample, other ages were well represented (3, 5, 6, 8, 9, 10, 12 and 13 years old). Secondary succession stands older than 14 years were not sampled due to the settlements' age. Since they were established between 1980 and 1984, and fieldwork was done in 1999, the uncertainty of sampling 15-year-old or older stands would be too high. Moreover, no landowner reported a fallow that old.
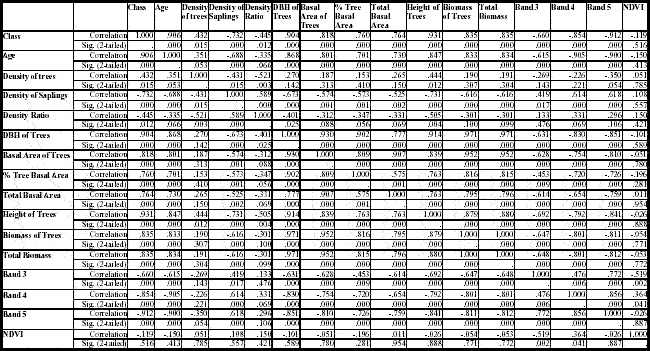
The sample variability allowed the comparison of vegetation structure and spectral responses within and across classes. In general, height and DBH of trees, density of saplings, total basal area, and total biomass were good indicators of vegetation regrowth stages. All of them were significantly separated among SS1, SS2, SS3, and forest classes (Table 9). It is important to mention that many of these variables are significantly correlated, indicating that less sampling effort would be needed to depict different classes of succession in broader studies at the regional scale (Table 11). For example, DBH, basal area, height, and biomass of trees are highly correlated. As other studies have shown, height or DBH of trees could be chosen in this case to represent the stage of regrowth (Moran et al. 2000). The advantage of choosing these variables instead of basal area or biomass is the relative simplicity of directly measuring them during fieldwork and perhaps in the future using Light Detection and Ranging (LIDAR) to estimate canopy height for large regional areas.
Table 11 - Pearson correlation coefficients for selected vegetation structure variables, mean reflectance in TM bands, and NDVI for sites sampled in Machadinho d’Oeste and Vale do Anari.
Despite the clear separation among classes of succession and forest, when graphed against mean reflectance in infrared TM bands (Figures 39 to 46), only three clusters of samples were well differentiated (SS1 and SS2 mixed together, SS3, and forest). These results supported the decision of grouping SS1 and SS2 into a single class of regrowth. In doing that, the accuracy increases in relation to the classification system. Also, the confusion between SS1 and pasture or SS2 and perennial agriculture tends to be minimized. In addition, two classes of succession are still maintained, allowing studies of land-cover dynamics through multi-temporal analyses. In sum, the decision of going from three to two classes of vegetation regrowth was necessary to improve the performance of further analyses of land-cover dynamics.
All these findings confirm the importance of land-use history besides age in defining stages of secondary succession. However, in more recent settlements such as Machadinho and Anari, age also may be significant because there was not enough time to produce the same impact as in older settlements. This is indicated by the significant correlation between age and vegetation structure variables or spectral data mentioned above (table 11). As pointed out by Uhl et al. (1988), site age is a good predictor of aboveground biomass accumulation on light- and moderate-use sites, but not on heavy-use sites. In this chapter, age and physiognomy were used as indicators for field sampling, but, after data collection, age also can become a variable to be analyzed together with vegetation structure variables and spectral responses in TM bands.
Although some studies have attempted to assign age to secondary succession stages based on total stand biomass (Nelson et al. 1999) or canopy geometry (Steininger 1996), what has been measured is the outcome of different trajectories of land use over distinct biophysical features. The stage of regrowth in terms of vegetation structure (and species composition), and not its age, is more useful in possible applications of mapping and monitoring succession classes. However, age may be appropriate as an organizing principle of regrowth stages when sampling vegetation and interviewing local people and landowners or to be used in studying the impact of land-use history on different sites (Moran et al. 2000). In this sense, knowledge of culture and context helps to achieve a better understanding of regeneration processes in the Amazon. Moreover, the study of vegetation structure variables such as total height of trees together with the investigation on biophysical features such as soil fertility and land-use history are better indicators regarding the degree of development of a regrowth stand. The assessment of these variables at local and regional scales and the interpretation of spectral data to depict such variability provide more accurate information about the trajectory of recovery occurring at distinct colonization stages in rural settlements in the Amazon.